经典回顾:肾脏类器官揭示TGFβ纤维化中SMAD3-EZH2协同调控染色质机制
来源:华津生物微信公众号
|
作者:华津生物
|
发布时间 :2026-05-27
|
45 次浏览:
|
🔊 点击朗读正文
❚❚
▶
|
分享到:
爱尔兰团队利用iPSC肾类器官结合单细胞RNA+ATAC测序,揭示TGFβ/SMAD3与EZH2在全基因组共占据。TGFβ刺激后,PDGFRA⁺成纤维细胞分化为ACTA2⁺/POSTN⁺肌成纤维细胞,伴随染色质大规模开放及增强子-启动子环连接。SCENIC分析鉴定出以SMAD3/AP-1为核心的“纤维化调节子”。EZH2抑制剂GSK343可阻断染色质开放、下调纤维化基因、减轻胶原沉积。研究揭示了SMAD3依赖EZH2酶活性开放染色质的机制,提出靶向二者表观遗传合作是抗纤维化新策略。
肾纤维化由TGFβ1驱动,其下游信号蛋白SMAD3入核后如何作用于染色质尚不清楚。已有研究发现SMAD3与负责关闭染色质的表观遗传酶EZH2之间存在相互作用,由此推测两者的协同合作可能是TGFβ信号通往染色质的关键环节。
为验证这一假说,2022年,爱尔兰都柏林大学Conway生物分子与生物医学研究所的John Crean教授团队在Communications Biology发表了题为"Single-cell multiomics reveals the complexity of TGFβ signalling to chromatin in iPSC-derived kidney organoids"的研究,利用iPSC来源的肾脏类器官建立纤维化模型,结合单细胞多组学技术,系统探究了这一表观遗传合作在纤维化中的作用,并测试了靶向该环节的治疗潜力。
提出新机制:揭示了TGFβ信号通过SMAD3与EZH2协同合作直接作用于染色质的新模型,发现两者在全基因组上共占据,SMAD3能识别并打开被EZH2锁闭的染色质区域。
技术整合创新:首次将单细胞RNA测序与单细胞ATAC测序联合应用,同步捕捉TGFβ1引起的转录组和染色质可及性变化,建立了从染色质开放到基因表达的完整调控链条。
发现核心调控网络:鉴定出驱动肌成纤维细胞分化的“纤维化调节子”,以SMAD3和AP-1为核心,包含ETS家族及NUAK1、CREB3L1、RARG等新发现的纤维化相关因子。
转化潜力明确:在人源iPSC来源的肾脏类器官中验证了EZH2抑制剂GSK343可阻断整个纤维化链条,为靶向SMAD3与EZH2表观遗传合作治疗肾纤维化提供了新策略。
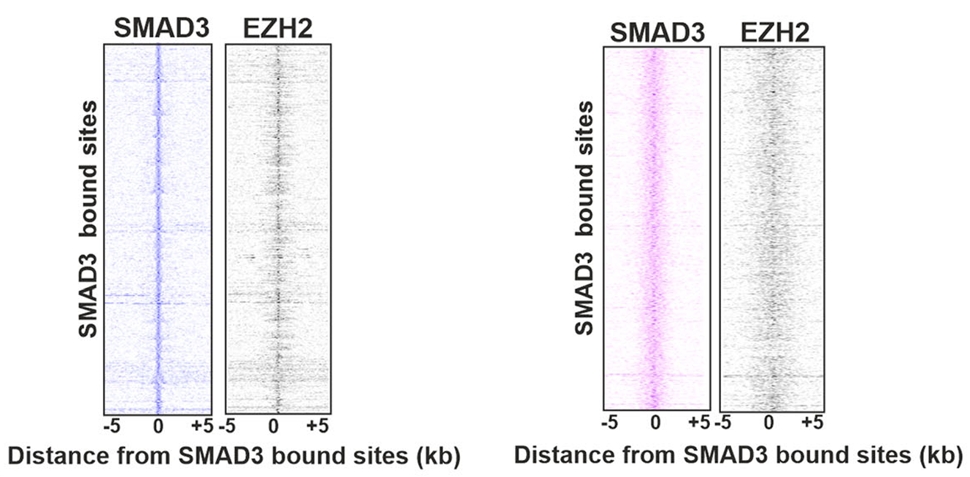
首先,研究团队用ChIP-seq技术扫描了全基因组,发现SMAD3和EZH2在iPSC和肾祖细胞(NPC)中大量共同占据在基因组的相同位置上,这种共定位为两者在功能上的协作提供了结构基础。
图1. SMAD3和EZH2在iPSC(图左)、NPC(图右)中共占据基因组。
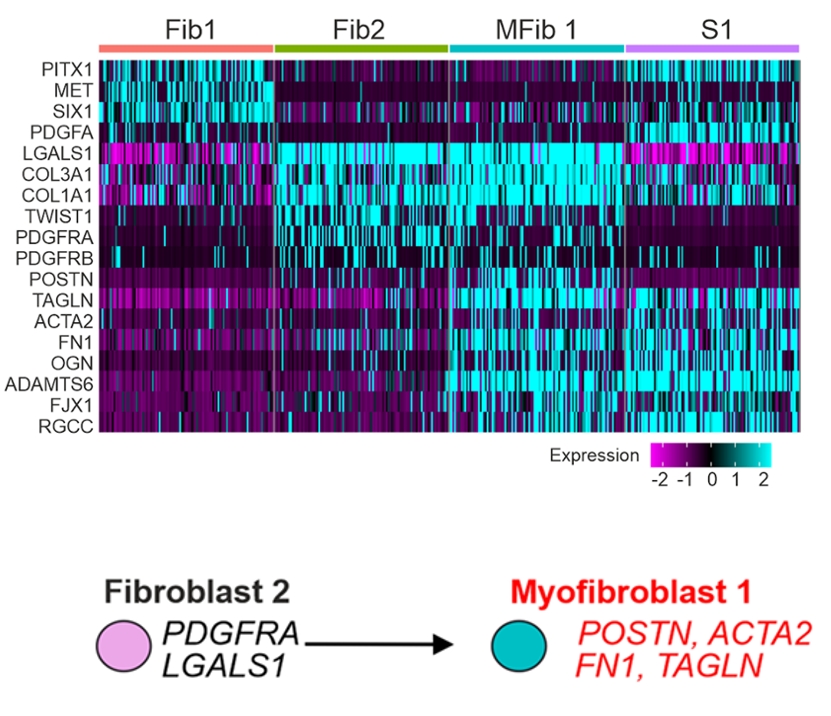
基于这一发现,研究者进一步利用人源iPSC培育出肾脏类器官,通过加入TGFβ1模拟体内纤维化环境,并结合单细胞RNA测序和ATAC测序追踪纤维化发生的全过程。结果显示,一群PDGFRA阳性的常驻成纤维细胞在TGFβ1的持续刺激下,分化为高表达ACTA2和POSTN的肌成纤维细胞。伴随这一转变,这些细胞的染色质经历了大规模重塑,大量纤维化相关基因的调控区域从紧闭变为开放。
图2. TGFβ1刺激后,PDGFRA⁺的Fib2(Fibroblast 1)分化为MFib1(Myofibroblast 1),两者基因表达谱的高度相似为这一谱系关系提供了分子证据。分化后的MFib1高表达ACTA2和POSTN,成为纤维化中产生瘢痕的主要效应细胞。
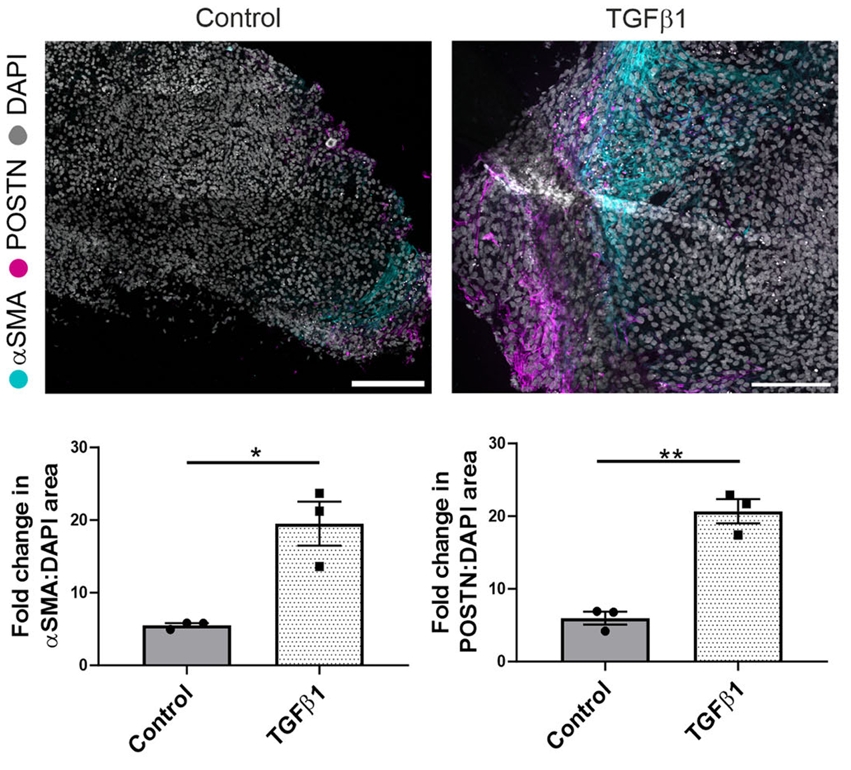
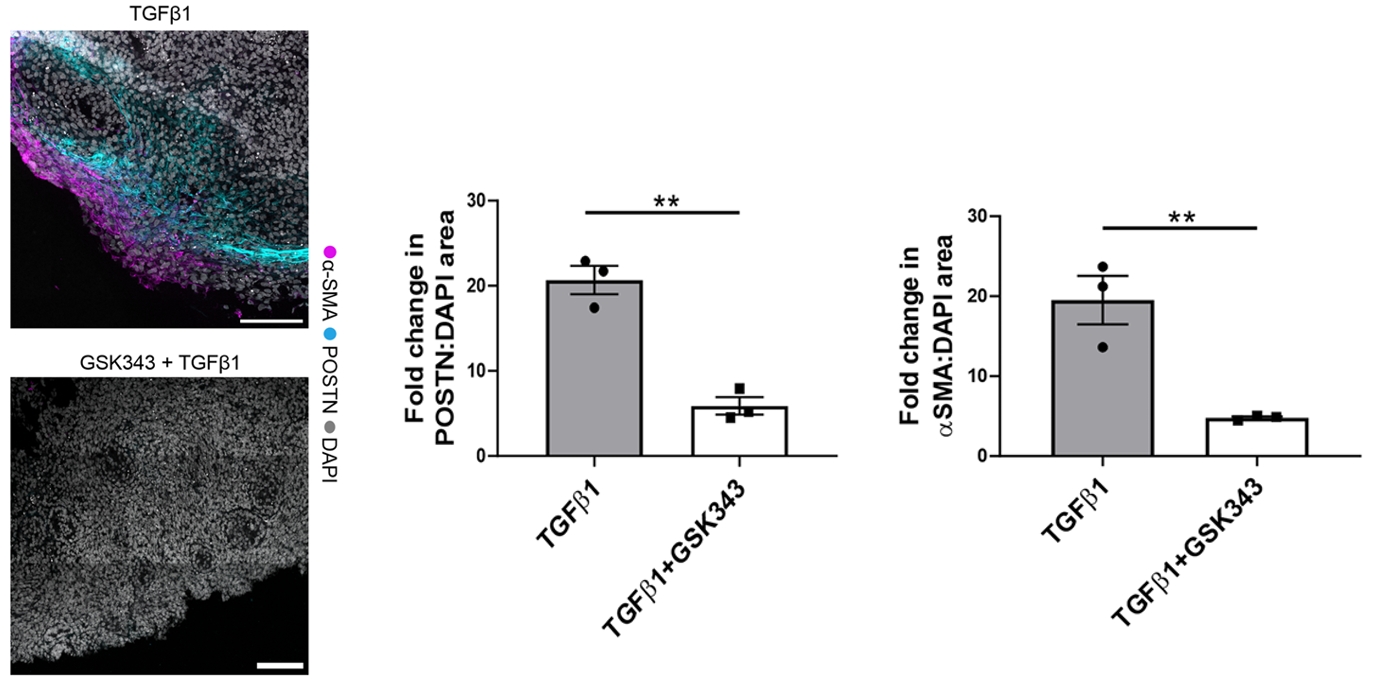
图3. 经TGFβ1处理后,肾脏类器官中α-SMA(由ACTA2基因编码)和POSTN的蛋白表达显著上调,从蛋白水平证实了前述PDGFRA阳性成纤维细胞向肌成纤维细胞的活化。
基于这些观察,研究者提出了一个机制模型:静息状态下,EZH2在纤维化基因上沉积抑制性标记,使染色质紧缩、基因沉默。当TGFβ信号激活SMAD3入核后,SMAD3识别这些被标记的位点,招募去甲基化酶和染色质重塑复合物,移除抑制标记、打开染色质,从而启动纤维化基因表达。
为验证这一模型,研究者用GSK343提前抑制EZH2的酶活性,再加入TGFβ1,(已知GSK343选择性抑制EZH2,不直接作用于SMAD3,且SMAD3与EZH2在基因组上预先存在共占据(图1)),结果发现,染色质不再开放(图6),增强子与启动子的互作被打断(图7),纤维化基因被显著削弱(图8),肌成纤维细胞分化也被抑制(图3)。这表明,在SMAD3激活和靶向识别未受直接干扰的情况下,仅阻断EZH2就足以使整条通路瘫痪。因此,SMAD3必须依赖EZH2的酶活性才能打开染色质,两者的协作是TGFβ信号通往染色质的必经闸门。
图4. 单独TGFβ1处理时,αSMA和POSTN的蛋白表达大幅上升;而用GSK343预处理后再给予TGFβ1,这两种蛋白的表达被显著压低。
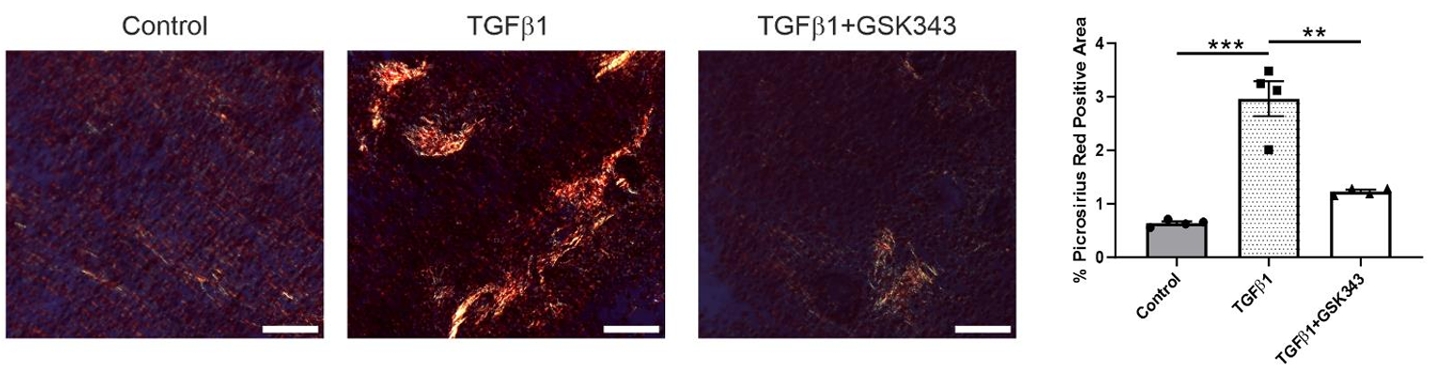
图5. 天狼星红染色显示,TGFβ1处理组胶原沉积较对照组显著增加,而GSK343预处理后胶原沉积较TGFβ1单独处理组显著减少,表明GSK343可有效减轻TGFβ1诱导的胶原积累。
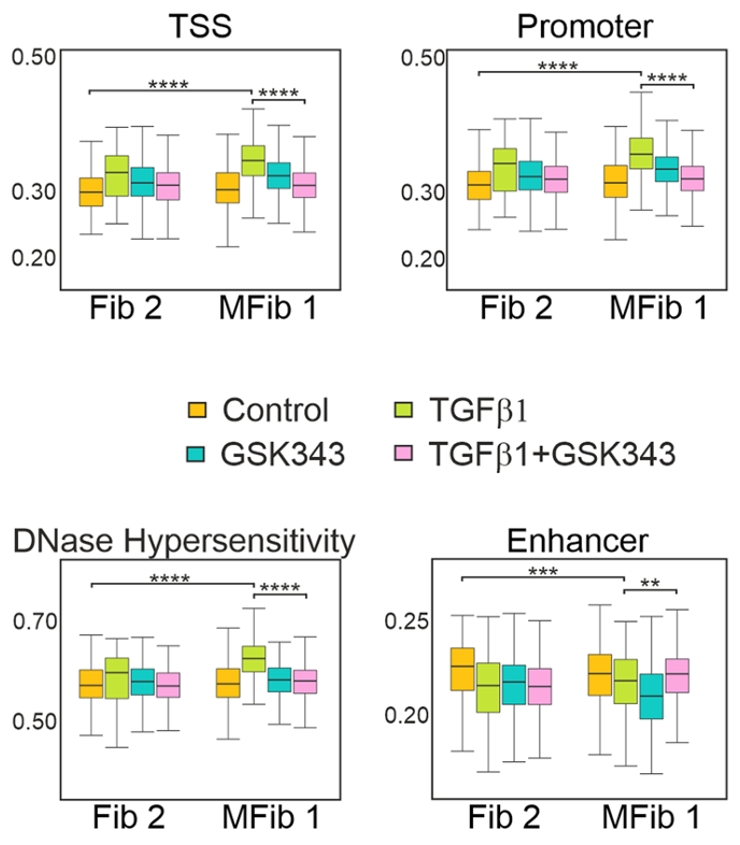
图6. GSK343可抑制TGFβ1引起的Fib2和MFib1中转录起始位点(TSS)、启动子(Promoter)、增强子(Enhancer)及DNase I超敏感位点(DNase I hypersensitivity)处的染色质可及性变化。
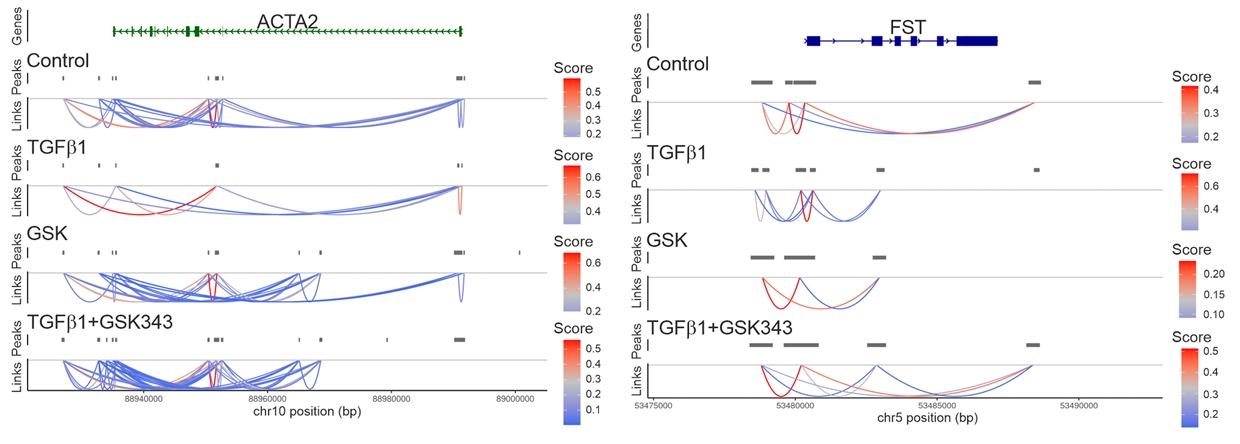
图7. TGFβ1能够增强纤维化关键基因上增强子与启动子之间的空间互作,而抑制EZH2后这种互作被逆转。
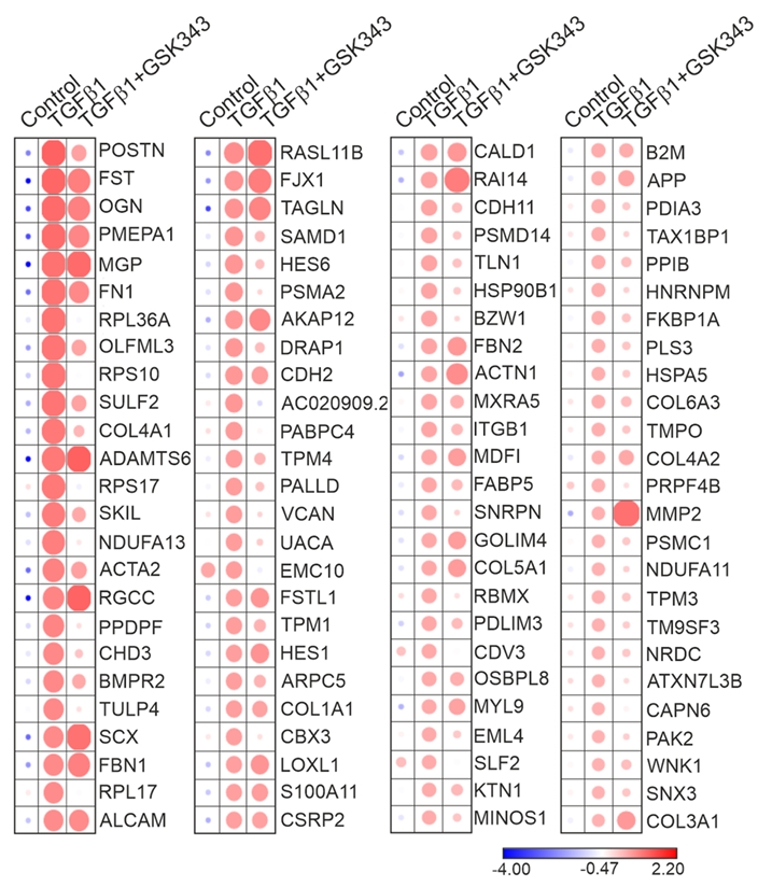
图8. GSK343抑制EZH2后,MFib1群体中TGFβ1诱导的纤维化相关基因表达被选择性下调。
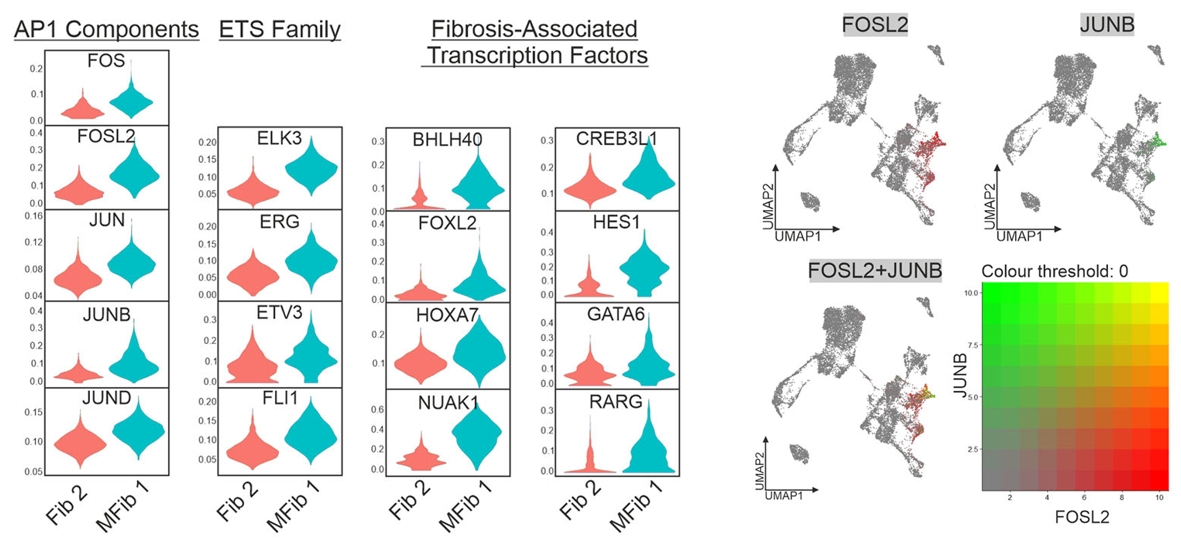
最后,研究者进一步追问了染色质打开之后执行纤维化程序的具体因子。通过SCENIC分析,他们鉴定出一个在肌成纤维细胞中被特异激活的转录因子调控网络,命名为“纤维化调节子”(“fibrotic” regulon)。该网络以SMAD3和AP-1家族成员(FOSL2、JUNB)为核心,同时包含ETS家族以及NUAK1、CREB3L1、RARG等新发现的纤维化相关因子,共同构成了驱动成纤维细胞向肌成纤维细胞终末分化的核心执行模块。
图9. “纤维化调节子”(含AP-1、ETS等)在MFib1中特异激活,FOSL2和JUNB的分布证实其驱动了肌成纤维细胞分化。
这项研究选用人类iPSC来源的肾脏类器官,主要基于两方面考虑。
在细胞组成层面,类器官含有多种肾脏细胞类型,具备与真实肾组织相似的细胞异质性和微环境,能够再现肾脏发育和疾病的复杂性。
在组织结构层面,它具有突出的自组织能力,可在培养中自发形成具有一定空间排布的三维结构,这一特性使其成为机制探究和药物干预测试的理想平台,研究者正是利用它来模拟TGFβ1诱导的纤维化反应,并在接近体内环境的条件下测试GSK343的干预效果。
研究者也坦诚指出该模型的局限性,即类器官整体偏向胚胎样组织,其中的足细胞仅处于不成熟前体阶段,裂孔隔膜标记物表达不明确。尽管如此,研究的所有核心发现均经过多模态技术的交叉验证,结论依然可靠。
这项研究利用iPSC来源的肾脏类器官,结合多模态单细胞分析,揭示了TGFβ1通过SMAD3与EZH2的协同合作来调控染色质可及性、激活纤维化特异性调节子、从而驱动肌成纤维细胞分化的完整分子机制。研究进一步证明,使用EZH2抑制剂GSK343可以阻断这一过程,减弱纤维化基因表达和染色质可及性变化。因此,靶向SMAD3与EZH2的关联,可能是治疗肾纤维化的一种有效策略。
参考文献:
Davis JL, Kennedy C, Clerkin S, Treacy NJ, Dodd T, Moss C, Murphy A, Brazil DP, Cagney G, Brougham DF, Murad R, Finlay D, Vuori K, Crean J. Single-cell multiomics reveals the complexity of TGFβ signalling to chromatin in iPSC-derived kidney organoids. Commun Biol. 2022 Nov 27;5(1):1301