近年来,肾类器官作为模拟肾脏发育和疾病的强大工具,已经广泛应用于药物筛选和疾病模型研究。传统上,人类肾脏类器官(由人类多能干细胞衍生)能够生成肾小管,但缺乏与收集管(CD)系统的连接,而这种连接是肾脏功能的基本构成部分。在今年的Cell Stem Cell上发表题为 “Integrating collecting systems in human kidney organoids through fusion of distal nephron to ureteric bud” 的研究展示了一种新颖的实验方法,不仅能在类器官中重建收集管网络,而且能够模拟真实的肾小管与输尿管芽的发育过程。该研究表明,GATA3等标志物在肾小管的远端区域表达,能够促进与输尿管芽的融合,从而完成肾脏功能关键部分的重建。
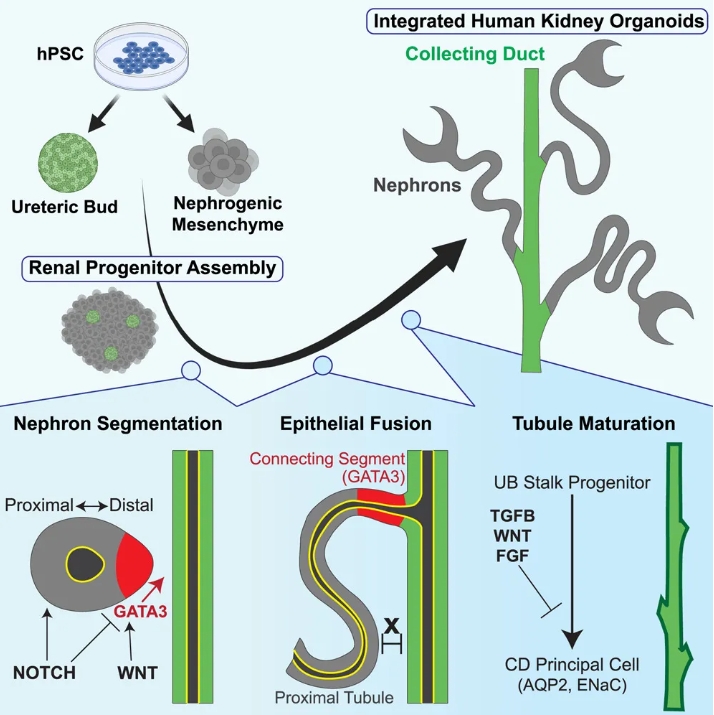
图形摘要
学术亮点
集合管形成:输尿管芽来源的球状体(UB spheroids)在肾脏类器官共培养中形成CD样管道,并与肾小管远端部分(通过GATA3+肾小管)成功融合。
GATA3表达:GATA3在肾小管远端区域的表达对于肾小管与收集管的融合至关重要。研究还发现,NOTCH和WNT通路调控了肾小管的远端模式化以及融合能力。
类器官成熟:通过调整信号环境,可以显著提高集合管和肾小管的成熟度,进一步推动类器官的生理功能化。
摘要
肾脏是人体重要的排毒器官,由数百万个肾单位(nephron)组成,这些肾单位通过远端与收集管(collecting duct, CD)系统融合,形成连续的管腔路径,用于排出代谢废物。然而,现有的基于人类多能干细胞(hPSC)的肾类器官模型虽能生成肾单位,但往往缺少完整的收集系统,导致肾小管以盲端形式终止,功能受限。这篇发表于2025年5月8日《Cell Stem Cell》杂志的论文,由美国辛辛那提儿童医学中心的Kyle W. McCracken团队领衔,题为Integrating collecting systems in human kidney organoids through fusion of distal nephron to ureteric bud。研究团队通过发育生物学启发的hPSC分化系统,成功实现了肾单位与输尿管芽(ureteric bud, UB)祖细胞的组装与融合,为构建功能性肾组织提供了新框架。
研究内容
1. UB-NM共培养体系的建立与细胞动态。
在类器官生成的初期,作者首先研究了如何通过将输尿管芽(UB)球体与肾源性间充质(NM)细胞进行结合,成功模拟出肾脏发育中的关键步骤。作者设计三步组装策略:将UB球体与NM共培养。5小时内NM围绕UB凝聚,2-4天UB延伸成管状上皮,NM分化为肾小泡(RV)。单细胞转录组证实:① UB(GFP+/GATA3+)与NM谱系严格分离;② UB保留分支潜能(RET+/WNT11+至第4天),但未能维持NPC生态位。此体系成功构建出UB管网与肾单位交织的类器官,为融合奠定结构基础。
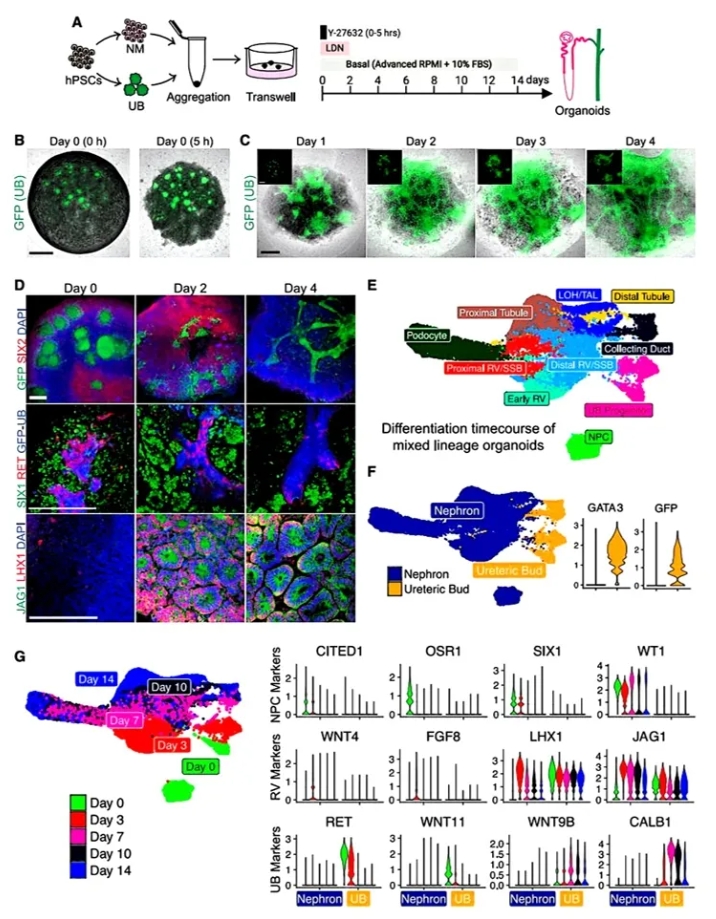
图1. UB和NM祖细胞组装成整合的肾脏类器官
2. UB spheroids与肾小管的融合。
在类器官生成后,作者继续探讨了UB spheroids与肾小管之间的融合过程。第14天单细胞聚类发现:混合类器官出现CD样细胞群(GFP+/CALB1+/AOP2+),而NM-only类器官缺失该群体。三维成像显示:GFP+ UB形成分支管网,其中96%的上皮连接点发生在GFP+收集管与GATA3+远端小管之间。而NM-only类器官的CALB1+连接段短且盲端,证实融合依赖远端身份特异性。时序追踪揭示:GATA3+区域主动延伸并"入侵"UB上皮,第5天完成顶膜连续性建立。这些结果表明,通过将UB spheroids与NM细胞共培养,可以有效促进收集管与肾小管之间的结构性整合。
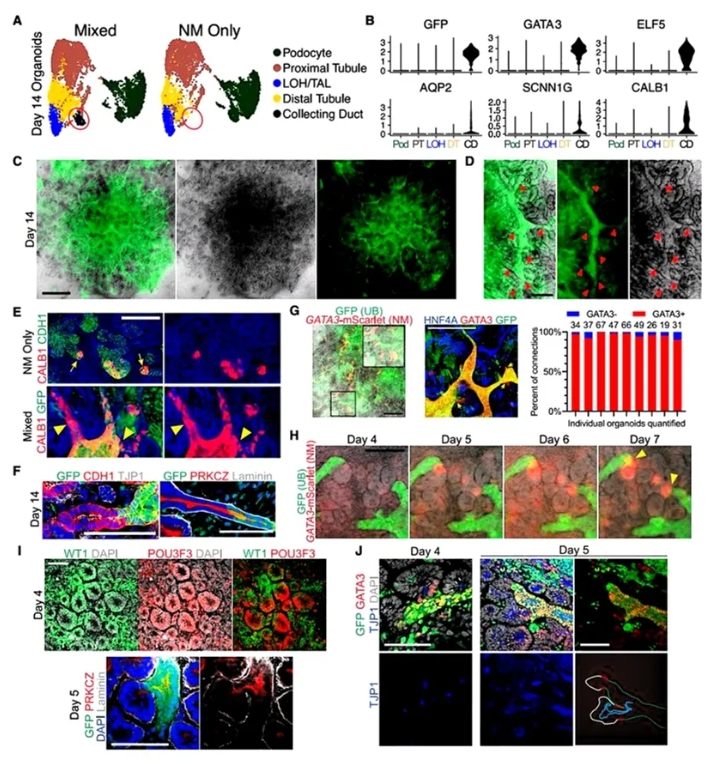
图2. 与远端肾单位小管融合的CDs的形成
3. 分段肾单位的成熟与UB诱导的远端偏移。
为明确UB对肾单位发育的影响,在UB与肾小管融合的基础上,作者进一步分析了不同信号通路对这一融合过程的调节作用。在第7天,类器官中的肾单位已经开始表现出明显的分段化特征。到第14天,肾单位的各个段落,如足细胞(NPHS1+)、近曲小管(LRP2+)和髓袢升支粗段(SLC12A1+)等,表现出了成熟的标志物。通过单细胞转录组分析,作者发现,混合类器官中的肾单位细胞群体表现出更高的成熟度,尤其是近曲小管和足细胞的分化。此外,混合类器官中的近曲小管和远端小管的比例显著增加,这一变化与UB的信号作用密切相关。总体来看,UB的加入对肾单位的远端化起到了促进作用。
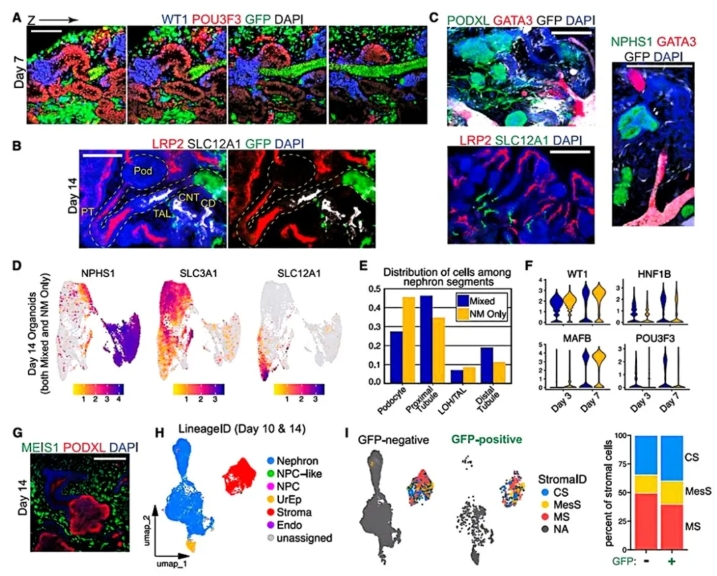
图3. 整合的类器官发育具有近端-远端极性的分段肾单位
4. 移植体内验证融合结构的生理稳定性。
为检验类器官体内功能性,作者将体外预融合的类器官移植至小鼠肾包膜下。2周后移植物血管化并形成分层肾实质:宿主血管灌注肾小球(含红细胞),NPHS1+足细胞与PDGFRB+系膜细胞有序排列。关键的是,GATA3+远端小管仍与长达1-2 mm的GFP+收集管融合。5周时收集管出现病理性扩张,暗示滤液潴留——这可能是类器官产生尿液的首个间接证据,但不幸的是,从这些结构中回收的流体体积太低,无法进行生化分析,无法明确评估它们是否反映了某种程度的肾功能或推定的尿液。
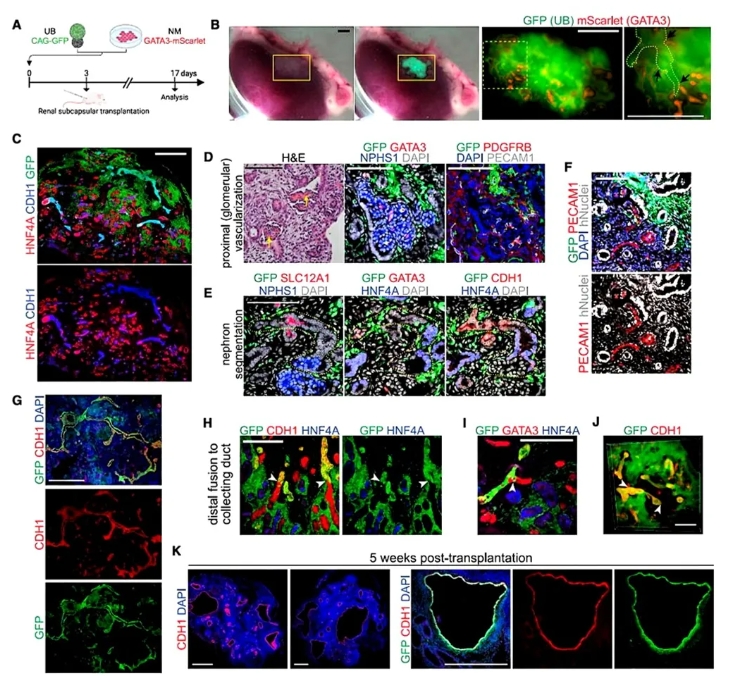
图4. 体内移植后UB与远端肾单位的融合
5. NOTCH抑制对融合效率的影响。
为进一步探讨肾单位与收集管融合的调节机制,作者分析了NOTCH信号对这一过程的影响。鉴于融合严格依赖GATA3+远端段,作者假设:融合频率可以通过改善GATA3+远端节段的规格来增加,重点是NOTCH操作是否可以用于调节近端。NOTCH信号抑制后,肾单位远端小管与输尿管的融合效率显著提高,导致GATA3+远端小管比例的增加,并且显著促进了肾单位与集尿管的融合。这表明,NOTCH信号通过影响肾单位的分化状态,调节了融合效率。NOTCH信号抑制可能是优化肾脏类器官融合的重要策略。
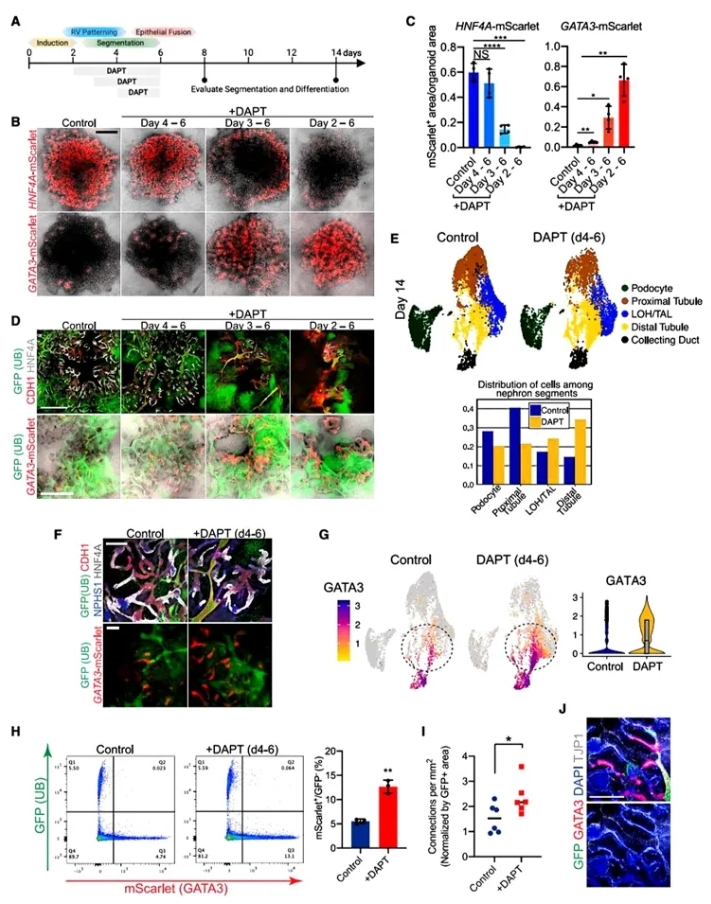
图5. NOTCH抑制增加远端肾单位规格和融合效率
6. WNT信号促进远端肾单位的融合。
为解析UB诱导远端偏移的机制,作者操控WNT通路:抑制(XAV939)降低GATA3并减少融合;激活(CHIR)则过度远端化。接着模拟UB微环境:在UB球体中诱导表达WNT9B,成功提升GATA3+远端段和融合频率。更关键的是,在WNT抑制背景下强制表达GATA3,可逆转融合缺陷,首次证明GATA3是融合的直接驱动因子。
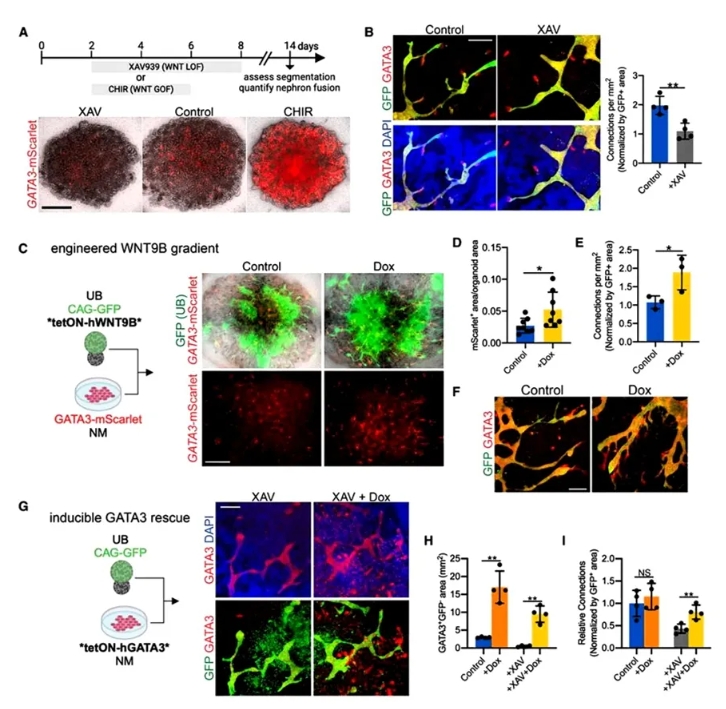
图6. WNT通过调节GATA3促进远端特异性和融合
7. 收集管成熟优化与跨平台比较。
最后作者提升CD功能:发现常规培养中CD细胞低表达成熟标志物AOP2。通过抑制WNT/TGF-β/MEK通路("AUX"方案),成功激活AOP2/SCNN1G等转运蛋白,构建出首个含AOP2+贯通CD的肾单位。横向对比4种类器官:Shi体系(本研究)的肾单位上皮占比最高(62.1%),且近端标志物SLC34A1和CD标志物AOP2表达最优,确立了新体系的成熟度优势。
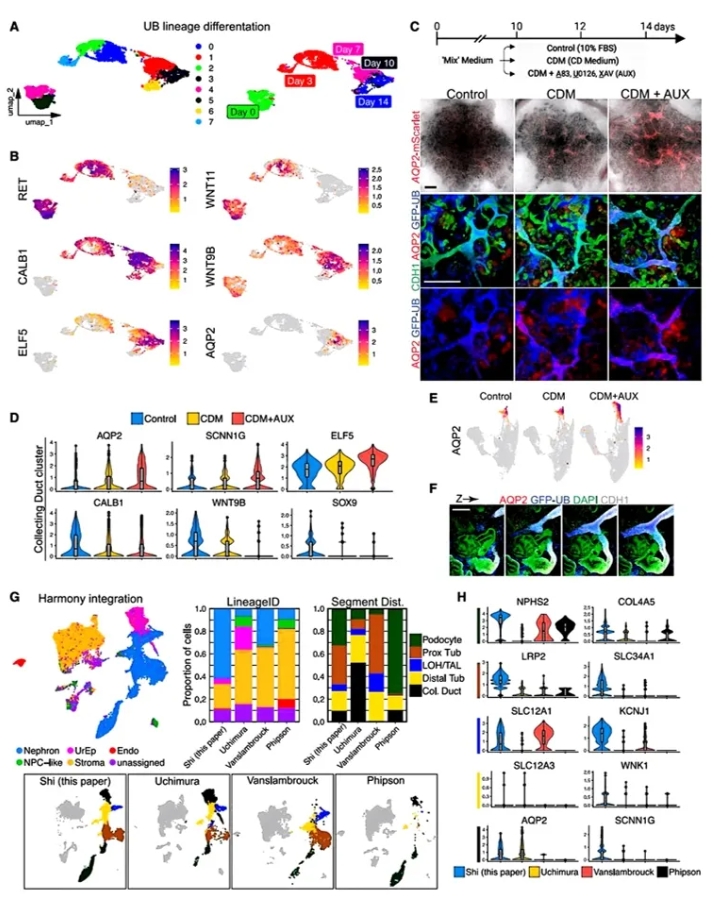
图7. 诱导重组类器官中CD成熟
总结
在结构上,这项研究首次在人肾类器官中显示了远端小管和UB之间的上皮融合的重演,这是形成肾单位极化和连接的关键事件,并且该系统在体外和体内移植后都是稳健的。从肾生物工程的角度来看,建立肾小管液从肾单位到收集系统的必要运动的途径是产生更多功能性肾组织的关键里程碑。此外,该系统为研究人类肾脏器官发生中肾单位融合的基本机制提供了前所未有的模型。通过这一系列的实验,作者不仅揭示了肾单位与输尿管融合的机制,还通过调节信号通路促进了肾脏类器官的成熟和功能化。这些发现为肾脏发育研究、疾病建模和再生医学提供了宝贵的实验平台和理论依据。
参考文献:
Shi M, Crouse B, Sundaram N, Pode Shakked N, Thorner K, King NM, Dutta P, Ester L, Zhang W, Govindarajah V, Kopan R, Cebrian C, Mayhew CN, Helmrath MA, Bonventre JV, McCracken KW. Integrating collecting systems in human kidney organoids through fusion of distal nephron to ureteric bud. Cell Stem Cell. 2025 Jul 3;32(7):1055-1070.e8.